\title {Philosophical Psychology \\ 10: What Are Those Motor Representations Doing There?}
\maketitle

10: What Are Those Motor Representations Doing There?
[email protected]
\def \ititle {10: What Are Those Motor Representations Doing There?}
\begin{center}
{\Large
\textbf{\ititle}
}
\iemail %
\end{center}
\section{The Double Life of Motor Representation}
\section{The Double Life of Motor Representation}
Motor representations live a kind of double life. Although paradigmatically involved in performing
actions, they also occur when merely observing others act and sometimes influence thoughts about the
goals of observed actions. Further, these influences are content-respecting: what you think about an
action sometimes depends in part on how that action is represented motorically in you.
- - - - - - - -
Suppose you are reaching for, grasping, transporting and then placing a pen. Performing even
relatively simple action sequences like this involves satisfying many constraints that cannot
normally be satisfied by explicit practical reasoning, especially if performance is to be rapid and
fluent. Rather, such performances require motor representations.
These representations are paradigmatically involved in preparing, executing and monitoring actions.%
\footnote{%
See \citet{wolpert:1995internal, miall:1996_forward, jeannerod:1998nbo, zhang:2007_planning}.
Note that motor representations sometimes occur in an agent who has prepared an action and is required (as it turns out) not to perform it: although she has prevented herself from acting, motor representations specifying the action persist, perhaps because they are necessary for monitoring whether prevention has succeeded \citep{bonini:2014_ventral}.
}
But they also live a double life. Motor representations concerning a particular type of action are
involved not only in performing an action of that type but also sometimes in observing one. That is,
if you were to observe Ayesha reach for, grasp, transport and then place a pen, motor representations
would occur in you much like those that would also occur in you if it were you---not Ayesha---who was
doing this.
Converging evidence for this assertion comes from a variety of methods and measures ...
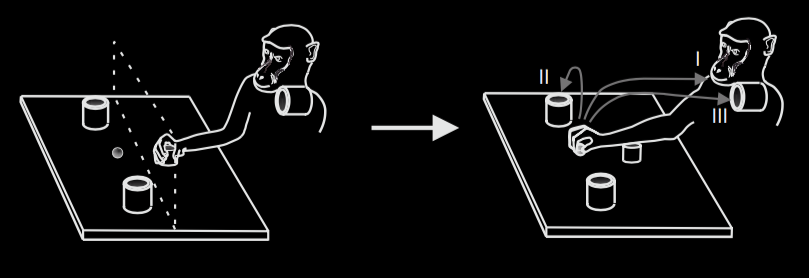
Fogassi et al 2005, figure 1B
Single cell recordings in nonhuman primates show that, for each of several types of action, there are
populations of neurons that discharge both when an action of this type is performed and when one is
observed \citep{pellegrino:1992_understanding, gallese:1996_action,Fogassi:2005nf}.
Fogassi et al 2005, figure 1B
This is the performance data.
Note that the neurons are firing before the distinctive part of the action has
occured: that is, the peak is between movement onset and the monkey first touching
the object to be grasped.
Now let’s compare
performance with observation.
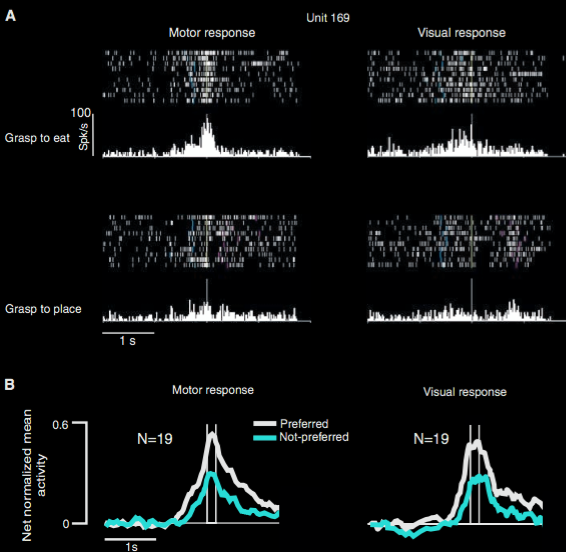
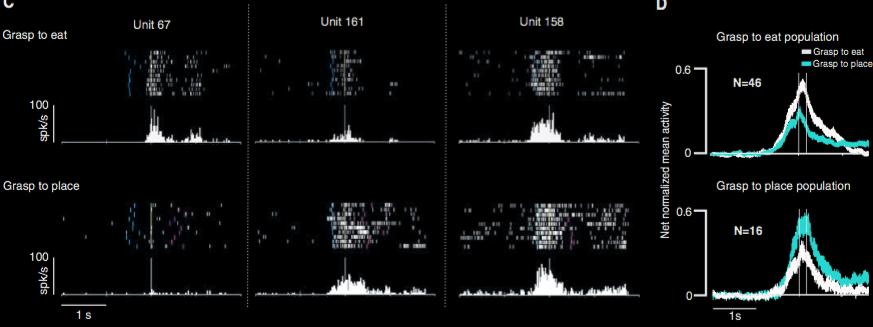
Fogassi et al 2005, figure 5
‘(A) Congruence between the visual and the motor response of a mirror neuron. Unit 169 has a stronger
discharge during grasping to eat than during grasping to place, both when the action is executed and
when it is observed. Conventions as in Fig. 1. (B) Population-averaged responses during motor and
visual tasks (12).’
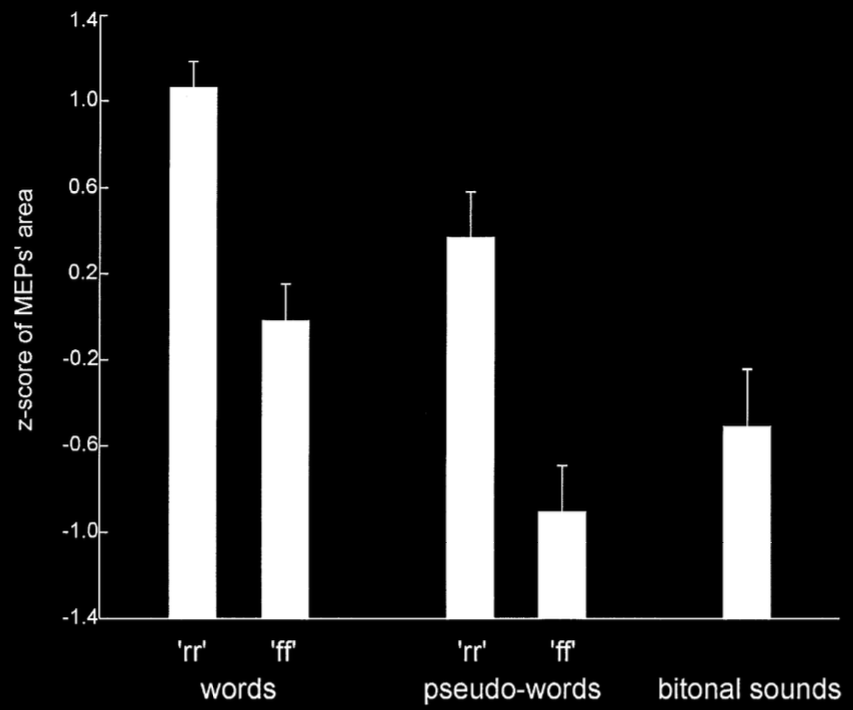
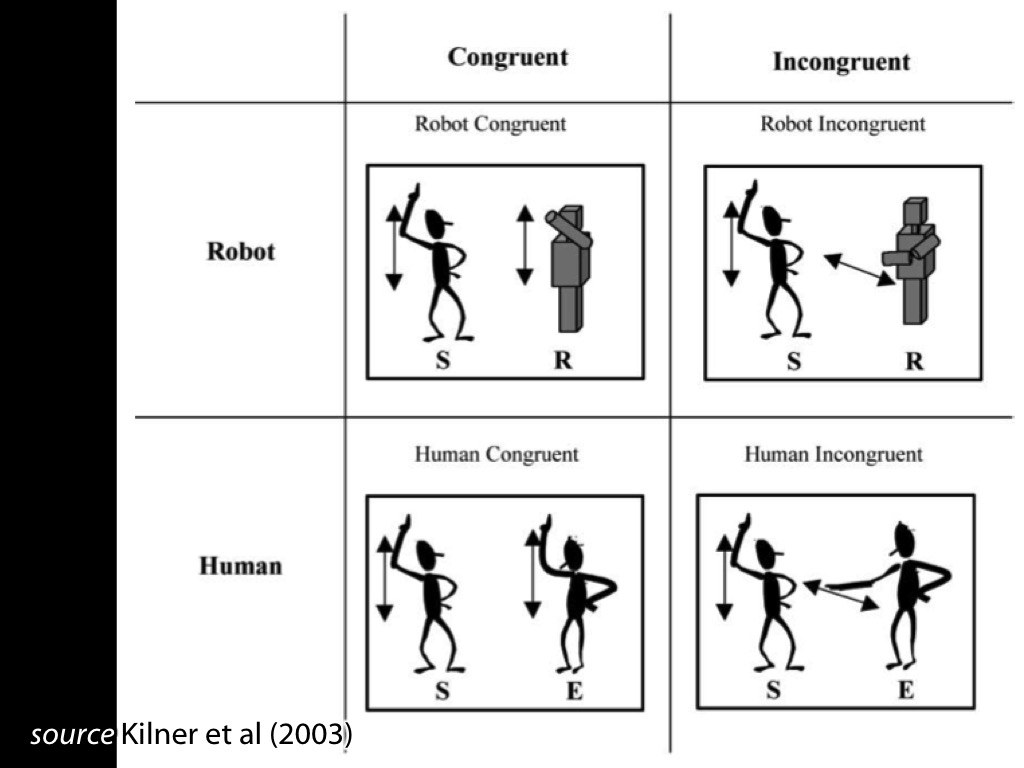
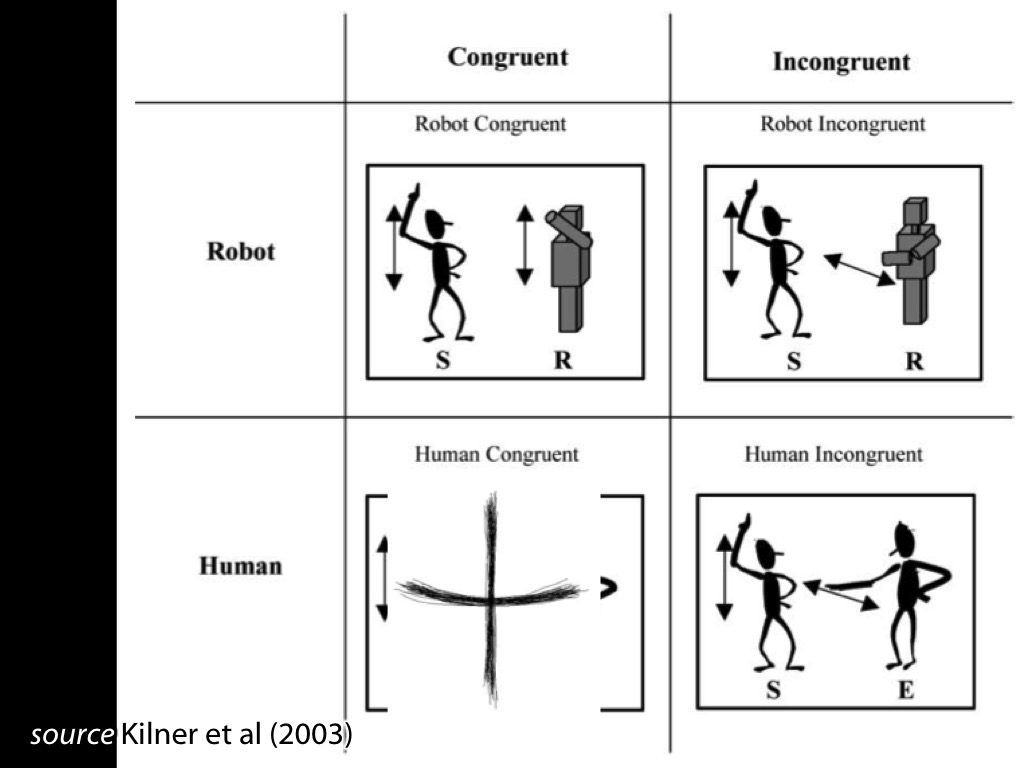
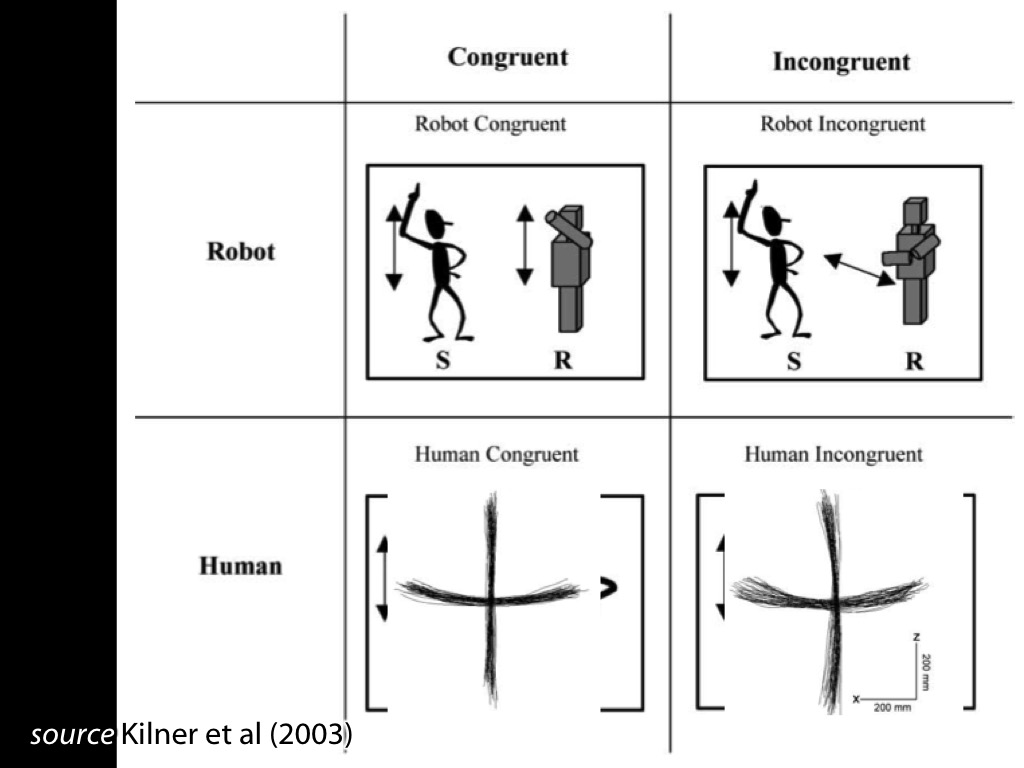
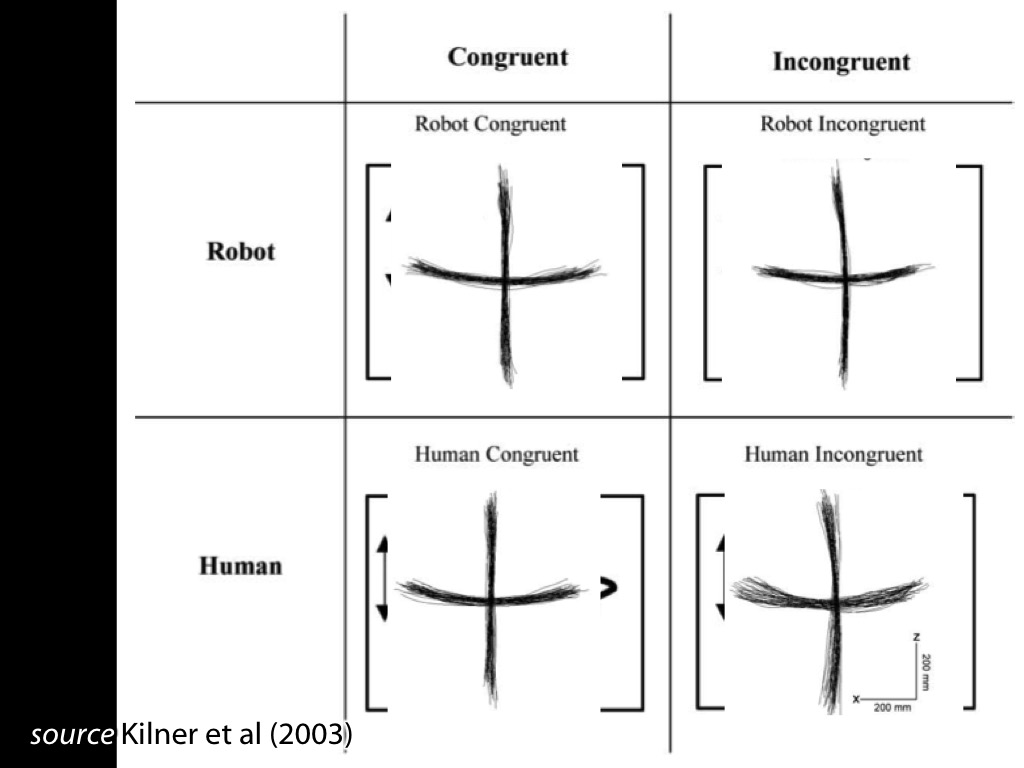
Fadiga et al 2002, figure 2

Behaviour: interference effects (ovalization)
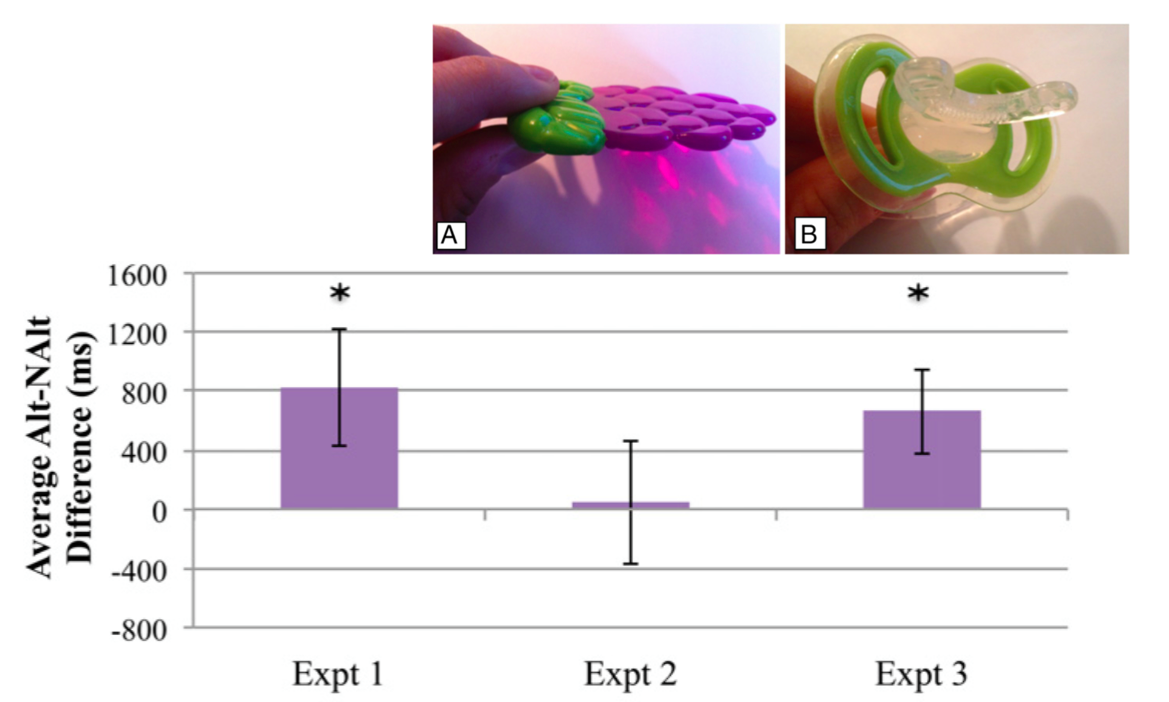
Bruderer et al, 2015 figures 1, 4
Experiment 1 : shows that 6-month-old infants can distinguish a phonetic contrast
they have never heard before (one that occurs in Hindi but not their linguistic
environments.) (The contrast used was the Hindi dental /d/̪ versus retroflex /ɖ/
distinction.)
These graphs show a difference in mean looking time between cases in which phonemes
are alternated and cases in which they are not. (Iff infants distinguish, they should
find the alternating phonemes more interesting.)
Experiment 2: but not when they have a tongue-controlling dummy in their mouths
Experiment 3: but yes when they have a dummy which leaves the tongue free.

Costantini et al, 2012
‘We recorded proactive eye movements while participants observed an actor grasping small or large
objects. The participants' right hand either freely rested on the table or held with a suitable grip
a large or a small object, respectively. Proactivity of gaze behaviour significantly decreased when
participants observed the actor reaching her target with a grip that was incompatible with respect to
that used by them to hold the object in their own hand.’
Follow ups: tie hands; TMS (impair)

Casile & Giese 2006, figure 1
Ability to perform actions: the 180 degree swing is standard walk, whereas
225 and 270 degree swings are not standard but can be trained.
Visual discrimination task: ‘The visual recognition experiment was based on a forced-choice
paradigm. In each trial, two point-light stimuli consisting of a total of nine dots were presented
successively, at two different positions on the screen ... Participants had to respond whether both
stimuli represented the same gait pattern. Four cycles of each gait pattern were presented, each
cycle lasting for about 1.2 s. The start position within the gait cycle was randomized across
trials.’
Then training while BLINDFOLDED.
Then visual recognition task again.
‘One possible explanation of the observed motor-visual transfer is that the
participants might have picked up the rhythm that characterizes the trained
motor pattern, but not necessarily the details of the learned body
movement. To rule out this possibility, we performed a control experiment
in which the motor training was replaced by purely visual training.’
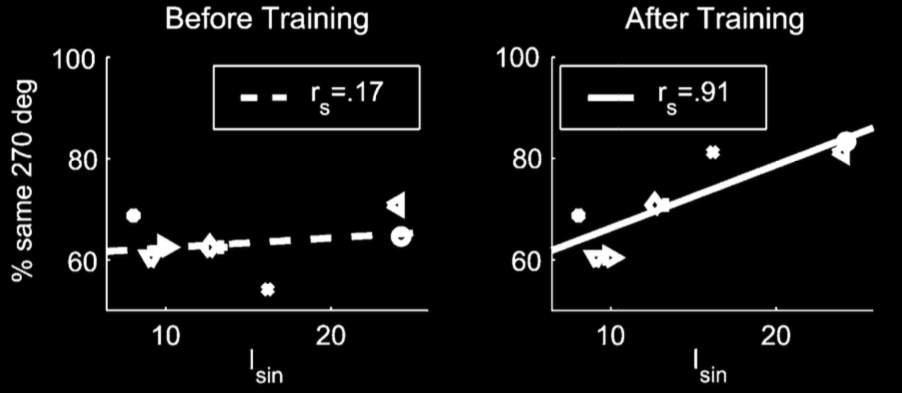
Casile & Giese 2006, figure 4A
Visual performance correlates with motor performance after (but not before) training.
(They also found no correlation with 225 degrees, which was not trained.)
This effect is perhaps surprising given that your judgement ultimately rests on purely visual
information (this is the point of the lights) whereas nothing could be seen during the training.
What explains this difference in judgement before and after training? Training of this kind typically alters the way things are represented motorically \citep{Calvo-Merino:2006ru}.
\label{expertise_affects_motor}
For this reason, the increase in the probability of making accurate judgements about the goals of others' actions is plausibly a consequence of differences in motor representations in the observer.

Pazzaglia et al 2007, figure 1B (part)
task: hear sound, identify one of four pictures.
Twenty-eight left-hemisphere-damaged patients with or without limb and/or buccofacial apraxia and
seven right- hemisphere-damaged patients with no apraxia were asked to match sounds evoking
human-related actions or nonhu- man action sounds with specific visual pictures.
‘In the novel sound-picture matching test used in this study, each patient was asked to listen to a
sound and then choose from among four pictures the one corresponding to the heard sound. The sounds
used included limb-related action sounds (LRAS), buccofacial-related action sounds (BRAS)
[ten sounds were transitive, i.e., object related (e.g., inflating a balloon) and ten were
intransitive, i.e., non object related (e.g., coughing). ], and non-human action-related sounds
(NHARS) [e.g. sea waves, breaking]’
Pazzaglia et al 2007, figure 1B (part)
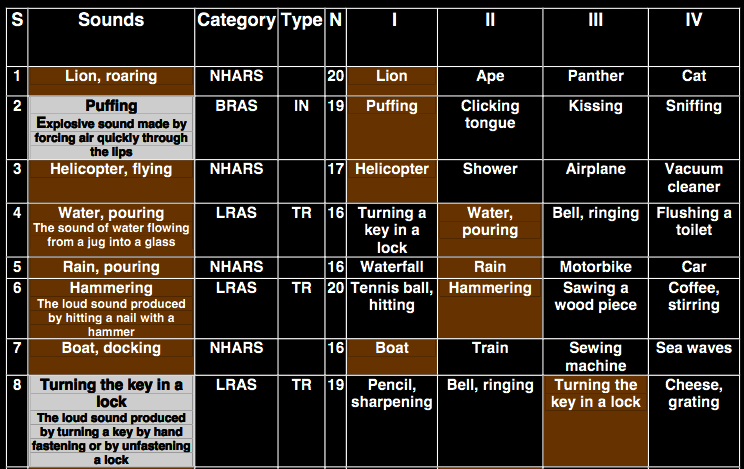
Pazzaglia et al 2007, Appendix S1 (fragment)
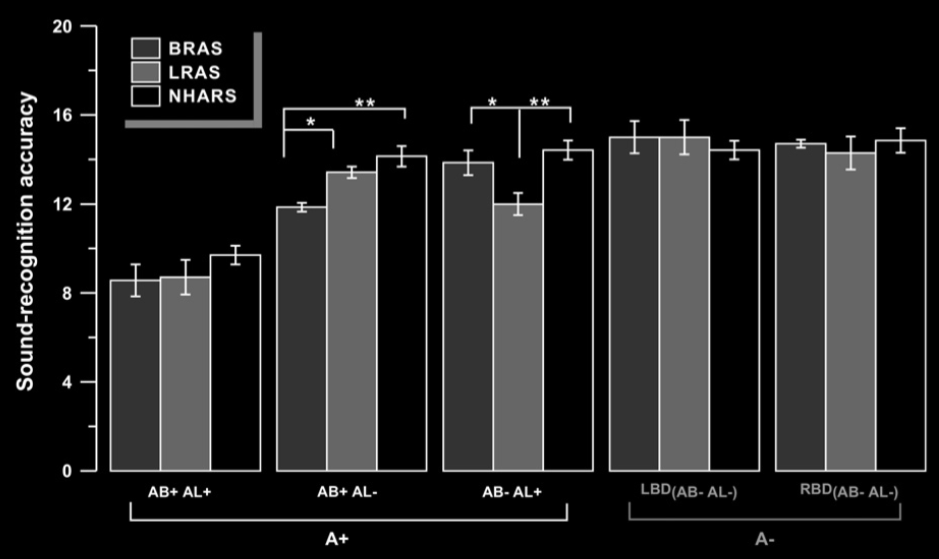
Pazzaglia et al 2007, figure 2
Beautiful results!
Key: limb-related action sounds (LRAS), buccofacial-related action sounds (BRAS), and non-
human action-related sounds (NHARS)
A+ apraxia; AB+ : buccofacial apraxia; AL+ limb apraxia; LBD: left brain damage; RBD: right brain damage
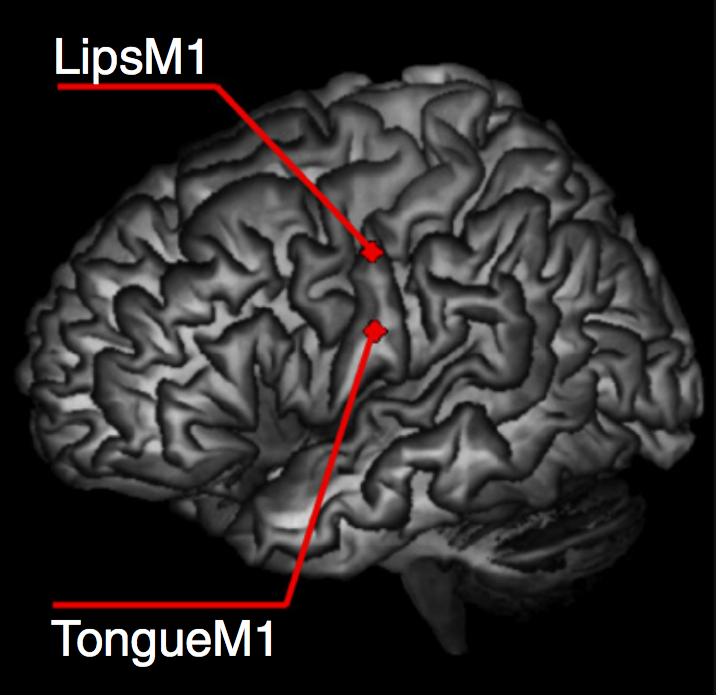
D'Ausilio et al (2009, figure 1)
‘Double TMS pulses were applied just prior to stimuli presentation to selectively prime the cortical activity specifically in the lip (LipM1) or tongue (TongueM1) area’
\citep[p.~381]{dausilio:2009_motor}
‘We hypothesized that focal stimulation would facilitate the perception of
the concordant phonemes ([d] and [t] with TMS to TongueM1), but that
there would be inhibition of perception of the discordant items
([b] and [p] in this case). Behavioral effects were measured via reaction
times (RTs) and error rates.’ \citep[p.~382]{dausilio:2009_motor}
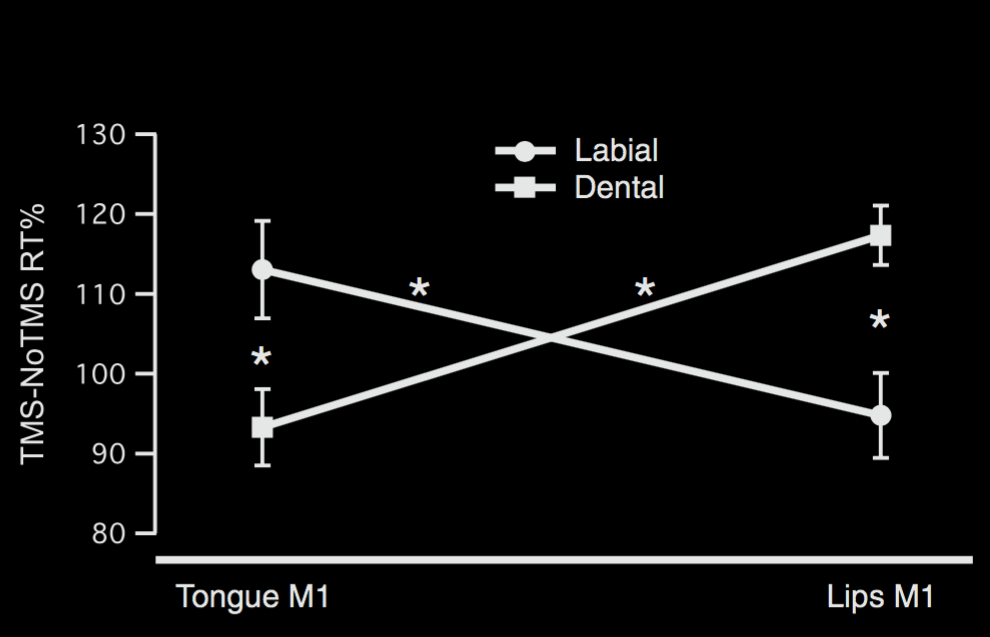
D'Ausilio et al (2009, figure 1)
‘Effect of TMS on RTs show a double dissociation between stimulation
site (TongueM1 and LipM1) and discrimination performance between class
of stimuli (dental and labial). The y axis represents the amount of RT
change induced by the TMS stimulation. Bars depict SEM. Asterisks
indicate significance (p < 0.05) at the post-hoc (Newman-Keuls) comparison.’
\citep{dausilio:2009_motor}